quinta-feira, 17 de outubro de 2024
domingo, 13 de outubro de 2024
Os erros inatos do metabolismo e a Genética
Erros Inatos do Metabolismo (EIM) são um grupo de doenças raras causadas por defeitos genéticos que afetam vias metabólicas essenciais no corpo humano. Esses distúrbios são, na maioria dos casos, herdados de forma autossômica recessiva e resultam de mutações em genes que codificam enzimas específicas ou proteínas envolvidas no metabolismo de carboidratos, proteínas, lipídios ou outros compostos bioquímicos. Essas mutações podem levar a uma produção inadequada ou à completa ausência da enzima, o que prejudica o funcionamento normal do organismo e pode causar o acúmulo de substâncias tóxicas ou a falta de compostos essenciais.
Principais tipos de Erros Inatos do Metabolismo
Os EIM são classificados de acordo com o tipo de substância que o corpo não consegue metabolizar adequadamente.
Algumas das principais categorias são
Distúrbios do metabolismo de carboidratos
Galactosemia: Deficiência na metabolização da galactose, um componente do açúcar do leite.
Doença de armazenamento de glicogênio: Afeta o armazenamento e a liberação de glicose.
Distúrbios do metabolismo de aminoácidos:
Fenilcetonúria (PKU): Incapacidade de metabolizar a fenilalanina, um aminoácido essencial.
Tirosinemia: Problemas na degradação do aminoácido tirosina.
Distúrbios do metabolismo de ácidos orgânicos:
Acidúria metilmalônica: Defeito no metabolismo dos ácidos orgânicos, causando acúmulo de ácido metilmalônico.
Acidúria propiônica: Deficiência na degradação de certos aminoácidos e lipídios.
Distúrbios do metabolismo de lipídios:
Doença de Gaucher: Deficiência de uma enzima que degrada glicocerebrosídeos.
Doença de Tay-Sachs: Problemas no metabolismo de gangliosídeos, um tipo de lipídio.
Distúrbios do ciclo da ureia:
Envolvem a incapacidade de eliminar adequadamente o amônio, um subproduto tóxico do metabolismo das proteínas. Exemplo: deficiência de ornitina transcarbamilase (OTC).
Distúrbios mitocondriais:
Relacionados à disfunção das mitocôndrias, organelas responsáveis pela produção de energia nas células. Exemplos: Síndrome de Leigh, deficiência de citocromo oxidase.
Genética dos Erros Inatos do Metabolismo
Os erros inatos do metabolismo são geralmente causados por mutações pontuais em genes específicos que codificam enzimas ou proteínas envolvidas em vias metabólicas. As mutações podem ocorrer de várias formas, como substituições de nucleotídeos, deleções ou inserções, e a gravidade da doença varia de acordo com o grau da deficiência enzimática.
Hereditariedade autossômica recessiva: A maioria dos EIM segue este padrão, o que significa que um indivíduo precisa herdar duas cópias defeituosas do gene (uma de cada pai) para desenvolver a doença. Se herdar apenas uma cópia, será portador, mas não manifestará sintomas clínicos.
Hereditariedade ligada ao X: Algumas doenças metabólicas, como a síndrome de Lesch-Nyhan, são causadas por mutações em genes localizados no cromossomo X e afetam principalmente os homens.
Expressividade variável e penetrância incompleta: Em algumas condições, a mutação genética não resulta necessariamente no desenvolvimento dos sintomas, ou pode haver variação na gravidade dos sintomas entre os indivíduos.
Diagnóstico
O diagnóstico dos EIM geralmente começa com a triagem neonatal, que inclui testes de sangue ou urina que detectam a presença de metabólitos anormais. Testes adicionais podem incluir:
Análise genética: Identificação de mutações específicas no DNA do paciente.
Espectrometria de massa: Usada para detectar anormalidades nos níveis de metabólitos.
Testes enzimáticos: Avaliação da atividade de enzimas específicas no sangue ou em tecidos.
Tratamento
O tratamento dos EIM varia conforme a doença e a gravidade. Algumas abordagens incluem:
Dieta restritiva: Para doenças como a fenilcetonúria, o controle da ingestão de alimentos que contêm o aminoácido problemático é essencial.
Suplementos enzimáticos: Alguns EIM podem ser tratados com suplementos que substituem as enzimas deficientes.
Terapia de reposição enzimática: Utilizada em doenças como a doença de Gaucher, onde enzimas específicas são administradas para substituir as que estão faltando ou são disfuncionais.
Terapia gênica: Uma abordagem mais recente que visa corrigir a mutação genética subjacente ao distúrbio.
Conclusão
Os erros inatos do metabolismo são condições genéticas que afetam o metabolismo, frequentemente diagnosticados na infância. Embora muitas dessas doenças ainda não tenham cura definitiva, o avanço em tratamentos genéticos e dietéticos tem melhorado significativamente o prognóstico de muitos pacientes. O diagnóstico precoce é crucial para evitar complicações graves e, em alguns casos, danos irreversíveis.
Aberrações Cromossômicas Estruturais
Aberrações cromossômicas estruturais referem-se a alterações na estrutura dos cromossomos que podem afetar a quantidade e a organização do material genético. Essas alterações podem ocorrer durante a divisão celular e podem ter consequências significativas para o organismo, podendo levar a doenças genéticas e câncer. As principais categorias de aberrações cromossômicas estruturais incluem:
1. Deleções
Descrição: Parte do cromossomo é perdida.
Consequências: Pode resultar em perda de genes importantes, levando a deficiências ou síndromes, como a síndrome de Cri du Chat.
2. Duplicações
Descrição: Segmentos do cromossomo são duplicados.
Consequências: Pode resultar em genes extras, o que pode levar a um aumento na produção de proteínas ou a doenças, como algumas formas de distrofia muscular.
3. Inversões
Descrição: Um segmento do cromossomo é invertido.
Consequências: Pode não causar problemas diretamente, mas pode afetar a segregação dos cromossomos durante a meiose, aumentando o risco de aneuploidias.
4. Translocações
Descrição: Segmentos de cromossomos são trocados entre cromossomos não homólogos.
Consequências: Podem ser equilibradas (sem perda de material genético) ou desequilibradas (com perda ou ganho de material). Translocações desequilibradas podem causar síndromes, como a síndrome de Down, quando envolvem o cromossomo 21.
5. Anéis Cromossômicos
Descrição: Formações em anel que ocorrem quando os extremos de um cromossomo se unem.
Consequências: Podem levar a problemas na divisão celular e estão associados a várias síndromes genéticas.
Causas das Aberrações Cromossômicas
Fatores Ambientais: Exposição a radiação, produtos químicos e infecções virais.
Erros na Replicação do DNA: Podem ocorrer durante a mitose ou meiose.
Fatores Genéticos: Algumas síndromes hereditárias podem predispor a aberrações.
Diagnóstico
Cito-genética: O estudo dos cromossomos é feito através da coloração e análise microscópica.
Técnicas Moleculares: Como o sequenciamento de DNA e a hibridização in situ (FISH), que ajudam a detectar e caracterizar aberrações.
Consequências Clínicas
As aberrações cromossômicas podem resultar em várias condições clínicas, incluindo:
Síndromes Genéticas: Como a síndrome de Turner, síndrome de Klinefelter e várias síndromes associadas a deleções ou duplicações.
Câncer: Muitas formas de câncer estão associadas a translocações e outras aberrações cromossômicas.
Conclusão
As aberrações cromossômicas estruturais têm um impacto significativo na biologia humana e na saúde. O entendimento dessas alterações é crucial para diagnósticos, tratamentos e aconselhamento genético, além de pesquisas sobre o desenvolvimento de doenças.
Aneuploidias Autossômicas
Aneuploidias autossômicas são um tipo de anomalia genética em que há uma alteração no número de cromossomos autossômicos, que são aqueles não relacionados aos cromossomos sexuais (X e Y). Em vez de ter o número normal de dois cromossomos de cada tipo (um de cada progenitor), uma pessoa com aneuploidia tem um número anormal de cromossomos. Isso pode resultar em problemas no desenvolvimento e em diversas síndromes. As aneuploidias autossômicas mais conhecidas envolvem a presença de um cromossomo extra (trissomia) ou a ausência de um cromossomo (monossomia).
Tipos comuns de aneuploidias autossômicas:
Trissomia do 21 (Síndrome de Down):
É a aneuploidia autossômica mais comum em seres humanos.
Caracteriza-se pela presença de uma cópia extra do cromossomo 21 (trissomia 21).
Sintomas: deficiência intelectual de grau variável, características faciais distintas, problemas cardíacos congênitos, entre outros.
A incidência aumenta com a idade materna.
Trissomia do 18 (Síndrome de Edwards):
Causada por uma cópia extra do cromossomo 18.
Sintomas: malformações graves, como defeitos cardíacos, retardo mental severo, orelhas de baixa implantação, e geralmente leva a uma expectativa de vida muito curta (a maioria dos afetados morre ainda na infância).
A síndrome é relativamente rara.
Trissomia do 13 (Síndrome de Patau):
Resulta da presença de uma cópia extra do cromossomo 13.
Sintomas: malformações severas do sistema nervoso central, defeitos cardíacos, defeitos craniofaciais (como lábio leporino), polidactilia (dedos extras) e retardo mental.
Assim como a Síndrome de Edwards, a expectativa de vida é curta, com muitos bebês falecendo nos primeiros meses de vida.
Síndrome de Cri du Chat:
Embora não seja uma aneuploidia no sentido clássico de trissomia ou monossomia, envolve uma deleção no cromossomo 5.
O nome da síndrome vem do choro característico dos bebês afetados, que se assemelha ao miado de um gato.
Sintomas: deficiência intelectual, microcefalia, e características faciais específicas.
Causas das aneuploidias:
A maioria das aneuploidias ocorre devido a erros na divisão celular durante a meiose, o processo que forma os gametas (óvulos e espermatozoides).
Esse erro é conhecido como não disjunção, quando os cromossomos não se separam corretamente, levando a gametas com um número incorreto de cromossomos. Se tal gameta anormal participa da fecundação, o embrião resultante terá uma aneuploidia.
Diagnóstico e Prevenção:
Diagnóstico Pré-Natal: Técnicas como a amniocentese e a biópsia de vilo corial podem detectar aneuploidias durante a gravidez. Há também exames de sangue mais recentes que analisam o DNA fetal circulante no sangue materno (NIPT – Teste Pré-Natal Não Invasivo).
Prevenção: Não há uma forma direta de prevenir as aneuploidias, mas fatores como a idade materna avançada aumentam o risco. Técnicas de reprodução assistida podem incluir testes genéticos pré-implantacionais para evitar a transferência de embriões aneuploides.
Consequências e Tratamento:
As aneuploidias autossômicas têm grande impacto no desenvolvimento físico e cognitivo. Não existe uma cura para essas condições, mas tratamentos sintomáticos e de suporte podem melhorar a qualidade de vida, especialmente em casos como a Síndrome de Down, onde as pessoas podem viver uma vida relativamente longa e saudável com os devidos cuidados médicos e suporte educacional.
Essas condições são bastante complexas e requerem suporte médico e familiar para garantir o melhor tratamento e acompanhamento possível.
Aneuploidias dos Cromossomos sexuais
As aneuploidias dos cromossomos sexuais são alterações numéricas envolvendo os cromossomos sexuais (X e Y), resultando em um número anormal de cromossomos sexuais em vez do par típico (XX para mulheres e XY para homens). Essas alterações podem impactar o desenvolvimento sexual, características físicas e, em alguns casos, funções cognitivas. As aneuploidias sexuais mais comuns inclue
1. Síndrome de Turner (45, X)
Cariótipo: Um único cromossomo X (45, X) ou, em casos raros, mosaico (45, X/46, XX).
Incidência: Cerca de 1 em 2.500 meninas.
Características: Mulheres afetadas geralmente têm baixa estatura, falha no desenvolvimento dos ovários (resultando em infertilidade), pescoço alado e problemas cardíacos. O desenvolvimento cognitivo é geralmente normal, mas algumas dificuldades de aprendizagem podem ocorrer.
2. Síndrome de Klinefelter (47, XXY)
Cariótipo: Um cromossomo X extra nos homens (47, XXY).
Incidência: Aproximadamente 1 em 500-1.000 nascimentos masculinos.
Características: Homens com Klinefelter têm testículos pequenos, baixa produção de testosterona, infertilidade, aumento do tecido mamário (ginecomastia) e, em alguns casos, dificuldades de aprendizagem ou linguagem. A maioria tem inteligência normal, mas pode haver dificuldades motoras.
3. Síndrome do Triplo X (47, XXX)
Cariótipo: Três cromossomos X (47, XXX) em mulheres.
Incidência: Cerca de 1 em 1.000 nascimentos femininos.
Características: Geralmente, as mulheres com esta síndrome têm uma aparência normal e fertilidade preservada. Algumas podem apresentar uma estatura mais alta do que a média e, ocasionalmente, atrasos no desenvolvimento motor ou dificuldades de aprendizagem, mas o impacto intelectual costuma ser leve ou inexistente.
4. Síndrome de Jacob ou XYY (47, XYY)
Cariótipo: Um cromossomo Y extra nos homens (47, XYY).
Incidência: Cerca de 1 em 1.000 nascimentos masculinos.
Características: Homens com 47, XYY geralmente são altos e podem ter um leve atraso na fala e aprendizado. A maioria tem fertilidade normal e desenvolvimento sexual típico, mas há relatos de problemas de comportamento ou dificuldades acadêmicas leves em alguns casos.
5. Mosaicismo de aneuploidias sexuais
Cariótipo: Algumas pessoas apresentam mosaicismo, onde diferentes linhas celulares têm diferentes números de cromossomos sexuais (por exemplo, 45, X/46, XX ou 45, X/46, XY).
Características: O fenótipo de indivíduos mosaicos depende da proporção das células normais e anormais. Alguns podem ter manifestações leves ou quase nenhuma, enquanto outros podem apresentar sintomas semelhantes aos de síndromes como Turner ou Klinefelter.
Diagnóstico e Tratamento
O diagnóstico dessas condições é feito por meio de testes genéticos, como o cariótipo ou testes de FISH (hibridização in situ fluorescente).
Tratamento: Focado em gerenciar os sintomas específicos. Pode incluir reposição hormonal (estrogênio ou testosterona), suporte educacional e, em alguns casos, intervenções cirúrgicas (como correção de ginecomastia ou tratamento de problemas cardíacos).
Essas aneuploidias demonstram como pequenas alterações no número de cromossomos podem levar a uma variedade de manifestações físicas, cognitivas e reprodutivas, destacando a importância dos cromossomos sexuais no desenvolvimento humano.
Aberraçãoes Cromossômicas
Aberrações cromossômicas são alterações no número ou na estrutura dos cromossomos, que podem ter impactos significativos no desenvolvimento, saúde e características dos organismos, especialmente em humanos. Estas aberrações podem ocorrer espontaneamente durante a divisão celular (meiose ou mitose) ou serem induzidas por fatores ambientais, como radiação ou exposição a certos agentes químicos. Elas são divididas em dois grandes grupos: numéricas e estruturais.
1. Aberrações Cromossômicas Numéricas
Essas alterações envolvem um número anormal de cromossomos, geralmente devido a erros na segregação cromossômica durante a meiose ou mitose. Os tipos principais incluem:
Aneuploidias: Presença de um número incorreto de cromossomos. Exemplos:
Síndrome de Down (Trissomia 21): Presença de três cópias do cromossomo 21.
Síndrome de Turner (Monossomia X): Presença de apenas um cromossomo X em mulheres.
Síndrome de Klinefelter (XXY): Presença de um cromossomo X extra em homens.
Poliploidias: A presença de um número de cromossomos que é múltiplo do conjunto normal, como em triploides (3n) ou tetraploides (4n). Esse tipo de aberração é mais comum em plantas, mas geralmente letal em humanos.
2. Aberrações Cromossômicas Estruturais
Essas aberrações ocorrem quando há modificações na estrutura de um ou mais cromossomos. Os principais tipos incluem:
Deleções: Uma parte do cromossomo é perdida. Exemplo: a Síndrome do Cri du Chat, causada pela deleção de uma porção do cromossomo 5.
Duplicações: Um segmento do cromossomo é duplicado, resultando em material genético extra.
Inversões: Um segmento do cromossomo é invertido, ou seja, ele é rearranjado de modo a estar em ordem inversa.
Translocações: Quando um segmento de um cromossomo é transferido para outro cromossomo não homólogo. Pode ser:
Recíproca: troca de segmentos entre dois cromossomos.
Robertsoniana: fusão de dois cromossomos acrocêntricos (com centrômero em posição terminal), resultando na perda de material genético.
Anéis Cromossômicos: Ocorre quando um cromossomo perde seus telômeros e as extremidades restantes se fundem, formando um anel.
Consequências das Aberrações Cromossômicas
Essas aberrações podem ter várias consequências:
Abortos espontâneos: Muitas aberrações cromossômicas graves resultam em perda precoce da gravidez, sendo uma das principais causas de abortos espontâneos.
Síndromes genéticas: Alterações cromossômicas podem resultar em síndromes genéticas com impactos no desenvolvimento físico e mental. Exemplos incluem as já mencionadas Síndromes de Down, Turner e Klinefelter.
Neoplasias: Em alguns casos, como em certos tipos de câncer, translocações cromossômicas específicas podem estar associadas à formação de tumores, como a leucemia mieloide crônica, que envolve uma translocação entre os cromossomos 9 e 22 (cromossomo Filadélfia).
Diagnóstico
O diagnóstico de aberrações cromossômicas pode ser feito por meio de cariótipos (análise microscópica dos cromossomos), FISH (hibridização fluorescente in situ) ou técnicas mais recentes como array CGH (hibridização genômica comparativa) e sequenciamento genético.
Considerações Finais
Aberrações cromossômicas são eventos biológicos que desempenham um papel crucial na genética médica. Seu estudo é essencial para compreender doenças genéticas, desenvolvimento de tratamentos e o aconselhamento genético para famílias que possuem histórico dessas condições.
Pleiotropia
A pleiotropia é um fenômeno genético no qual um único gene afeta múltiplas características fenotípicas aparentemente não relacionadas. Em outras palavras, um gene pleiotrópico pode influenciar vários aspectos do desenvolvimento e da fisiologia de um organismo. Esse conceito é importante na genética porque mostra como a função de um único gene pode ter impactos amplos no organismo como um todo.
Exemplos de Pleiotropia:
Síndrome de Marfan: Um exemplo clássico de pleiotropia ocorre na Síndrome de Marfan, uma doença causada por mutações no gene FBN1, responsável pela produção da fibrilina, uma proteína do tecido conjuntivo. As pessoas com essa mutação podem apresentar diversas manifestações, como problemas cardíacos, esqueléticos e oculares.
Anemia Falciforme: Outro exemplo é a anemia falciforme, que resulta de uma mutação no gene HBB que codifica a hemoglobina. Essa mutação provoca a deformação das hemácias, levando a uma série de sintomas, como anemia, dor e problemas nos órgãos, como o baço e os rins, mostrando múltiplos efeitos de um único gene.
Fenilcetonúria (PKU): É uma condição genética em que a mutação do gene PAH afeta a capacidade do organismo de metabolizar o aminoácido fenilalanina, o que pode levar a uma variedade de problemas neurológicos e cognitivos, além de outros sintomas metabólicos.
Mecanismo da Pleiotropia
A pleiotropia ocorre porque os genes codificam proteínas ou enzimas que podem estar envolvidas em múltiplos processos biológicos diferentes. Por exemplo, uma enzima pode ser crucial tanto no desenvolvimento de estruturas físicas quanto no metabolismo, e sua mutação pode causar defeitos em ambas as áreas.
Relevância Biológica e Evolutiva
A pleiotropia tem um papel significativo na evolução, pois uma mutação que beneficia uma característica pode ter efeitos adversos em outras, criando um equilíbrio entre seleção natural e efeitos colaterais. Isso faz com que as mudanças evolutivas dependam de como o gene afeta múltiplas características ao mesmo tempo.
Considerações
A pleiotropia também é importante em medicina, pois ajuda a entender como algumas mutações genéticas podem causar um conjunto de sintomas diversos, o que é crucial para o diagnóstico e tratamento de doenças genéticas complexas.
Esse conceito amplia a compreensão de que os genes não agem isoladamente e podem ter efeitos amplos e interconectados em um organismo.
Herança Quantitativa
Herança quantitativa é um tipo de herança genética em que uma característica fenotípica é controlada por vários genes (poligenia), e não por um único gene, como ocorre na herança mendeliana simples. Isso resulta em uma distribuição contínua dos fenótipos em uma população, em vez de categorias discretas. Ou seja, os fenótipos variam em grau ao longo de um espectro, como altura, cor da pele, peso e inteligência.
Aqui estão alguns pontos-chave sobre a herança quantitativa:
Poligenia: Características quantitativas são influenciadas por muitos genes (chamados de loci poligênicos), onde cada gene tem um pequeno efeito sobre o fenótipo total. Esses genes atuam de forma aditiva, ou seja, o efeito total no fenótipo resulta da soma dos efeitos individuais de cada gene.
Efeito ambiental: Além dos fatores genéticos, o ambiente também desempenha um papel importante na determinação das características quantitativas. Por exemplo, a nutrição pode afetar a altura de uma pessoa, além da contribuição genética.
Distribuição contínua: Diferentemente das características mendelianas, que resultam em poucos fenótipos distintos, as características quantitativas apresentam uma variação contínua. Um exemplo clássico é a altura em humanos, que não se restringe a poucas categorias, mas sim varia de forma gradual entre os indivíduos, formando uma curva de distribuição normal (em forma de sino).
Conceito de variância: Na herança quantitativa, a variação total observada no fenótipo de uma população pode ser decomposta em variância genética (VG), variância ambiental (VE) e, às vezes, variância genética-ambiental (VGE). A relação entre a variância genética e a variância fenotípica total é chamada de herdabilidade.
Exemplo clássico: cor da pele: A cor da pele em humanos é um exemplo comum de herança quantitativa. Ela é determinada pela interação de vários genes, com cada um contribuindo para a produção de melanina. A combinação de muitos alelos leva a uma variação contínua de tons de pele.
Curva de distribuição normal: Como muitas características quantitativas resultam da ação de muitos genes, a soma de seus efeitos tende a produzir uma distribuição normal (curva em forma de sino) na população, como no caso de altura, peso e pressão arterial.
Estudo de características quantitativas
No estudo de herança quantitativa, os geneticistas usam ferramentas estatísticas, como a análise de variância (ANOVA) e métodos para estimar a herdabilidade. Esses métodos permitem entender quanto da variação fenotípica observada se deve aos genes e quanto se deve ao ambiente.
Aplicações
A herança quantitativa é crucial em áreas como a genética de populações e o melhoramento genético, como na agricultura, onde características como rendimento de culturas e resistência a pragas são influenciadas por muitos genes.
Também é estudada na genética médica, especialmente em relação a doenças complexas, como diabetes, hipertensão e distúrbios mentais, que são influenciadas por múltiplos genes e fatores ambientais.
Resumo
A herança quantitativa trata de características complexas que dependem de múltiplos fatores genéticos e ambientais, com a variação fenotípica contínua entre os indivíduos. Isso a diferencia da herança mendeliana simples, onde um gene define diretamente o fenótipo.
Interação de Genes Não Alelos
A interação de genes não alelos refere-se ao fenômeno em que genes localizados em diferentes loci (posições no cromossomo) interagem entre si para influenciar a expressão de uma característica. Esses genes, que não são alelos entre si, podem modificar o efeito de outros genes, levando a variações fenotípicas não previstas por herança mendeliana simples. As principais formas de interação entre genes não alelos incluem:
1. Epistasia
A epistasia ocorre quando um gene interfere na expressão de outro gene em um locus diferente. O gene que mascara o efeito de outro é chamado de epistático, enquanto o gene cujo efeito é mascarado é chamado de hipostático.
Existem diferentes tipos de epistasia:
Epistasia dominante: Um alelo dominante em um locus mascara a expressão de alelos em outro locus. Exemplo clássico ocorre em plantas de cor branca, onde um gene dominante "suprime" o gene que define a cor.
Epistasia recessiva: Um alelo recessivo em um locus impede a expressão de um gene em outro. Por exemplo, em camundongos, um gene recessivo pode bloquear a produção de pigmento, levando a uma cor branca, independentemente do gene que definiria a cor do pelo.
2. Complementação
A complementação acontece quando dois genes não alelos precisam interagir para produzir um fenótipo específico. Cada gene contribui com uma parte essencial do processo. Se qualquer um dos genes for mutante, o fenótipo normal não se manifesta.
Um exemplo disso ocorre em flores com cores específicas. Dois genes diferentes podem codificar enzimas em uma via biossintética para a produção de pigmento. Se ambos funcionarem, a cor será expressa, mas se um deles falhar, a cor não será formada.
3. Herança poligênica
Na herança poligênica, várias pares de genes contribuem cumulativamente para a expressão de uma característica. Esses genes podem estar em loci diferentes, e o efeito de cada gene é aditivo. Características como altura, cor da pele e peso são exemplos clássicos de herança poligênica.
4. Gene modificador
Genes não alelos podem atuar como modificadores, alterando a intensidade ou a expressão de um gene em outro locus. Esse tipo de interação pode suavizar ou intensificar o fenótipo causado pelo gene principal.
5. Pleiotropia
Embora a pleiotropia não seja uma interação entre genes diferentes, vale mencionar que um único gene pode afetar várias características fenotípicas. Isso ocorre porque um gene pode ter múltiplas funções ou estar envolvido em diferentes vias biológicas.
Exemplos práticos:
Cor da plumagem em galinhas: A cor da plumagem em galinhas é um exemplo clássico de epistasia recessiva. O gene que determina a produção de pigmento (B) pode ser mascarado por um gene epistático recessivo (b), resultando em galinhas brancas, mesmo que a galinha possua os genes para uma plumagem colorida.
Metabolismo em humanos: Em humanos, condições como a fenilcetonúria (PKU) podem ser causadas pela interação de múltiplos genes não alelos que regulam o metabolismo de aminoácidos.
Conclusão
A interação de genes não alelos é fundamental para entender a variabilidade genética observada em organismos. Através de mecanismos como epistasia e complementação, essas interações resultam em um conjunto mais complexo de padrões hereditários do que o previsto pelas leis mendelianas tradicionais. Esses processos são essenciais em genética e biologia molecular para explicar a diversidade fenotípica observada nas populações.
Epistasia
A epistasia é um fenômeno genético que ocorre quando um gene interfere ou modifica a expressão de outro gene. Nesse contexto, dizemos que um gene é epistático (o que faz a supressão) e o outro é hipostático (cuja expressão é alterada).
Ela faz parte de interações entre genes não alélicos, ou seja, genes que estão em diferentes loci (posições no cromossomo). A epistasia pode ocorrer tanto em genes que codificam características específicas quanto em genes que regulam funções mais gerais, como o desenvolvimento.
Tipos de epistasia:
Epistasia dominante: Ocorre quando a presença de um alelo dominante em um gene impede a expressão de um outro gene, mesmo que esse outro gene também possua alelos dominantes. Um exemplo clássico envolve a cor da pelagem de certos animais, como cães e camundongos.
Epistasia recessiva: Aqui, dois alelos recessivos de um gene bloqueiam a expressão de um outro gene. O exemplo mais clássico é o sistema de cor da pelagem em camundongos, onde um gene recessivo pode "esconder" a cor da pelagem, resultando em uma pelagem albina.
Epistasia duplicada: Quando dois genes diferentes têm o mesmo efeito fenotípico e, independentemente da combinação de seus alelos, ambos acabam levando ao mesmo fenótipo. Isso pode ser comum em sistemas redundantes no organismo, onde múltiplos caminhos bioquímicos podem levar a um resultado semelhante.
Epistasia aditiva: Os efeitos de múltiplos genes podem se somar, levando a uma interação complexa em que a expressão de um gene é influenciada pela soma dos efeitos de vários outros genes.
Exemplo clássico de epistasia:
Um exemplo bem conhecido envolve a coloração da pelagem dos cães labradores. Dois genes diferentes controlam a cor do pelo: um controla a produção de pigmento (B/b, onde B é preto e b é marrom), e o outro controla a deposição do pigmento no pelo (E/e, onde E permite a deposição e e impede). Assim, cães com o genótipo ee terão pelagem amarela, independentemente do que está presente no gene para cor.
Importância da epistasia:
A epistasia é importante na genética porque ajuda a explicar a complexidade de como os traços são herdados e expressos, indo além do simples conceito mendeliano de dominância e recessividade. Ela é particularmente relevante em estudos de herança quantitativa, como em caracteres poligênicos, onde muitos genes podem estar envolvidos na determinação de um único traço.
Além disso, a epistasia também é importante para a genômica e estudos de doenças genéticas, pois algumas interações entre genes podem predispor ou proteger os indivíduos contra certas condições.
LINKAGE
O linkage é um termo utilizado em diversas áreas, sendo particularmente importante em estatística, biologia (genética) e análise de dados. Vou apresentar os principais usos desse termo nas diferentes áreas:
1. Linkage em Genética
No contexto da genética, linkage refere-se ao fenômeno em que dois ou mais genes estão localizados próximos no mesmo cromossomo e tendem a ser herdados juntos. Quando os genes são fisicamente próximos, a probabilidade de que sejam separados durante o processo de recombinação meiótica é menor, o que significa que eles serão herdados como uma unidade.
Mapa de Linkage: É uma representação que mostra a ordem dos genes e as distâncias entre eles no cromossomo, medido em unidades de recombinação. Isso ajuda a prever quais características genéticas serão transmitidas juntas.
Linkage Desequilíbrio: Trata-se de uma associação não aleatória de alelos em diferentes loci, indicando que eles são herdados juntos com maior frequência do que seria esperado por acaso.
2. Linkage em Estatística e Aprendizado de Máquina
Na estatística, especialmente na análise de agrupamentos (clustering), linkage é uma técnica usada para medir a similaridade entre clusters. É amplamente utilizada nos algoritmos de hierarchical clustering (agrupamento hierárquico) para decidir como agrupar pontos de dados.
Existem vários tipos de linkage usados na análise hierárquica:
Single Linkage (Ligação simples): Mede a distância entre os pontos mais próximos de dois clusters. Esse método tende a produzir clusters alongados.
Complete Linkage (Ligação completa): Mede a distância entre os pontos mais distantes de dois clusters. Gera clusters mais compactos.
Average Linkage (Ligação média): Calcula a média das distâncias entre todos os pares de pontos nos dois clusters.
Ward’s Linkage: Minimiza a soma dos quadrados das diferenças dentro de cada cluster, buscando otimizar a variabilidade intra-grupo.
Cada um desses métodos influencia a maneira como os dados são agrupados e a forma final dos clusters.
3. Linkage em Mecânica (Sistemas de Barras)
Em mecânica, o termo linkage se refere a mecanismos que consistem em barras rígidas conectadas por articulações móveis. Esses mecanismos são usados para transmitir movimento ou força. Um exemplo clássico de linkage é o mecanismo de quatro barras, que converte movimento rotacional em movimento linear ou vice-versa.
Linkage cinemático: Refere-se a sistemas em que várias partes estão ligadas para realizar movimentos desejados.
Exemplos: São comumente usados em robótica, automóveis (suspensões), e em várias máquinas industriais.
A Relação da Meiose e a Segunda Lei de Mendel.
A Segunda Lei de Mendel, também conhecida como Lei da Segregação Independente, afirma que os alelos de diferentes genes segregam de forma independente uns dos outros durante a formação dos gametas. Isso significa que a herança de um par de alelos não afeta a herança de outro par de alelos, desde que os genes envolvidos estejam em cromossomos diferentes (não ligados).
Essa lei está diretamente relacionada ao processo de meiose, que é a divisão celular que dá origem aos gametas (espermatozoides e óvulos) em organismos sexuados. Para entender essa relação, é importante considerar o comportamento dos cromossomos durante a meiose.
2°LEI DE MENDEL

MEIOSE
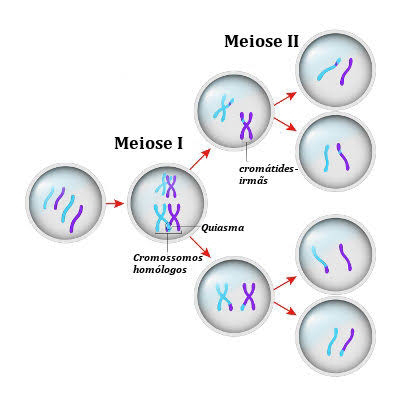
Relação entre a Segunda Lei de Mendel e a Meiose:
Meiose e Segregação dos Alelos: Durante a meiose, os pares de cromossomos homólogos (que possuem os mesmos genes, mas podem ter alelos diferentes) se alinham no plano equatorial da célula e são segregados aleatoriamente para diferentes células-filhas durante a meiose I. Cada gameta recebe um cromossomo de cada par, ou seja, um alelo de cada gene.
Essa segregação dos cromossomos homólogos é o que fundamenta a primeira lei de Mendel (lei da segregação), já que alelos de um mesmo gene (presentes em cromossomos homólogos) são separados durante a formação dos gametas.
Segregação Independente dos Genes:
A segunda lei de Mendel ocorre porque, durante a meiose I, os pares de cromossomos homólogos se alinham de forma independente na placa metafásica. Assim, a separação de um par de cromossomos (e seus alelos) é independente da separação de outro par.
Isso implica que a combinação de alelos para dois genes diferentes será distribuída de forma independente para os gametas, resultando em uma variedade de combinações genéticas.
Crossing-over e Exceções:
O processo de crossing-over (ou permutação), que ocorre na prófase I da meiose, pode gerar uma mistura adicional de material genético, aumentando a variabilidade genética. No entanto, o crossing-over pode ocorrer entre genes que estão próximos no mesmo cromossomo, o que pode levar à recombinação e não à segregação completamente independente, o que é uma exceção à segunda lei de Mendel. Esses genes são chamados de ligados.
Exemplo Prático:
Mendel estudou a segregação de dois genes que influenciavam a cor e a textura das sementes de ervilhas. Ele observou que o alelo para semente amarela (A) segregava de forma independente do alelo para textura lisa (B), resultando em várias combinações nos descendentes: sementes amarelas e lisas, amarelas e rugosas, verdes e lisas, verdes e rugosas. Esse comportamento corresponde ao que acontece durante a meiose: os cromossomos que contêm esses genes se separam independentemente.
Conclusão:
A meiose é o processo biológico que permite a separação e combinação independente dos alelos, como descrito na segunda lei de Mendel. A segregação dos cromossomos homólogos durante a meiose I garante que os alelos de genes localizados em cromossomos diferentes sejam herdados independentemente.
A Segunda Lei de Mendel.
A Segunda Lei de Mendel, também conhecida como a Lei da Segregação Independente, foi proposta por Gregor Mendel em seus estudos sobre a herança de características em plantas, especialmente em ervilhas. Essa lei descreve como diferentes pares de alelos segregam (se separam) de maneira independente uns dos outros durante a formação dos gametas. Aqui estão os principais pontos sobre essa lei:

Fundamentos da Segunda Lei de Mendel
Segregação Independente: Mendel observou que, quando se cruzam organismos que diferem em duas ou mais características, cada par de alelos se distribui nos gametas de forma independente. Isso significa que a herança de um caráter não afeta a herança de outro.
Experimentos com Ervilhas: Mendel conduziu experimentos com ervilhas em várias características, como cor da semente e forma da semente. Ao cruzar plantas que eram híbridas para diferentes características, ele notou que as características se combinavam de forma previsível na próxima geração.
Genótipos e Fenótipos: A partir dos cruzamentos, Mendel estabeleceu que as combinações de alelos determinam os fenótipos observáveis (características físicas) dos descendentes. Os fenótipos são o resultado da combinação dos alelos que os indivíduos herdam de seus progenitores.
Exemplo Prático: Se considerarmos duas características, como cor da semente (amarela e verde) e forma da semente (lisa e rugosa), a combinação dos alelos de cada característica ocorre de forma independente. Portanto, os descendentes podem exibir qualquer combinação dessas características.
Diagrama de Punnett: Para ilustrar a Segunda Lei de Mendel, os geneticistas usam o diagrama de Punnett, que ajuda a prever os genótipos e fenótipos possíveis dos descendentes a partir de cruzamentos de pais com diferentes características.
Importância da Segunda Lei
Base da Genética: A Segunda Lei de Mendel é fundamental para a compreensão da genética moderna. Ela estabelece as bases para o entendimento da variabilidade genética e da herança de características em organismos.
Herança Multifatorial: Essa lei é especialmente relevante em estudos de herança multifatorial, onde múltiplos genes influenciam características complexas.
Avanços em Melhoramento Genético: A compreensão da segregação independente permitiu avanços em áreas como agricultura, medicina e biotecnologia, ao possibilitar a previsão de como características específicas podem ser herdadas em organismos.
Limitações
Embora a Segunda Lei de Mendel tenha sido revolucionária, também possui limitações:
Ligação Genética: Mendel não conhecia o conceito de ligação genética, onde genes localizados próximos uns dos outros em um cromossomo tendem a ser herdados juntos, o que pode violar a segregação independente.
Interação entre Genes: A Segunda Lei não leva em conta interações entre diferentes genes, que podem influenciar os fenótipos de maneiras mais complexas do que a simples segregação independente.
Conclusão
A Segunda Lei de Mendel é um pilar fundamental da genética que ajudou a moldar nossa compreensão da herança e da variabilidade genética. Os conceitos introduzidos por Mendel continuam a ser aplicados em pesquisas modernas e em práticas de melhoramento genético.
Assinar:
Comentários (Atom)